行動変化がもたらす間接的な種間関係
松田裕之(東大海洋研)
まつだひろゆき
生態系は常に個体数が変動する非定常系である。個体数の変化は生物の行動や形質の変化を誘い、形質変化は他種の環境を変える。このような形質変化は間接的な種間関係をもたらし、しばしば直感に反する影響を及ぼす。
巻き添え競争と間接効果
同じ餌を利用する2種の捕食者は、互いに競争関係にある。一方の捕食者の個体数が増えると、被食圧が増えて餌生物の数が減り、他方の捕食者の摂食量が減る。これを取り合い競争または消費型競争(exploitative competition)という。[]
これと同様に、共通の天敵に狙われる2種の被食者は、互いに競争関係にある。一方の被食者の個体数が増えると、それを利用する捕食者が増え、他方の被食者への被食圧も高まる。これを巻き添え競争(apparent competition)という。[]この場合、2種の被食者は直接餌や住み場所などを巡って争うわけではない。天敵がいて初めて2種の間に関係が生じる。このように、第3の種を介して生じる種間関係を間接効果(indirect effect)という。[]
間接効果の最も典型例が巻き添え競争である。この概念が提唱されたのは1977年にことである。20年を経た今日、ようやく巻き添え競争の実証研究が報告され始めた。[]
群集内のそれぞれの種の個体数は他種との関係によって時間的に変化する。つまり、生態系は相互作用によって動的に変化する力学系である。しかし、生物は個体数やその空間分布、齢構造だけでなく、行動(behavior)や形質(trait)、すなわち表現型(phenotype)も変化する。さらに個々の生物の表現型によって、他種に与える影響は変わる。ではその行動や形質がどのように変化するかと言えば、自分の子孫の数を増やすように変わると考えられる。これを形質の適応的変化(adaptive change)と言い、それを研究する分野が進化生態学(evolutionary ecology)である。[]
形質の適応的変化を考慮すれば、間接効果も違って見えてくる。これを形質変化を通じた間接効果(trait-mediated indirect effect)という。[]
餌選好と被食者の共存
形質変化を考慮すれば、共通の天敵に狙われる2種の被食者は、必ずしも競争関係にあるとは限らない。もしも捕食者が2種の被食者を満遍なく狙うのでなく、多い方を集中して狙うとすれば、別の結果を導くことがある。
一般に、環境中の被食者の種組成(相対密度)と捕食者の胃内容物中の種組成は異なる。ある被食者iの環境中の頻度xiより胃の中の頻度yiが高いとき、捕食者はその餌を選り好んでいると考えられる。餌選好性を表す指数としてはShoryginの指数Ei=(yi/
?yj)/(xi/?xj)とそれを応用したIvlevの指数(Ei-1)/(Ei+1)が古くから使われていた。Eiが1より大きいとき、つまりIvlevの指数が正のときにはその餌が選り好みされていることを表す。けれども、Ivlev指数はほとんど利用しない餌を含めてEiを計算するかどうかで符号や値が変わるという欠点があった。J. Chessonの指数(yi/xi)/Σ(yj/xj)はこの欠点を克服したものである。[]胃の中の餌組成は環境中の餌組成に左右される。常にある餌種が好まれているなら胃の中の組成はそれほど影響を受けないが、好んでいた餌が環境中に少ないときには別の餌を食べるようになる。環境中の頻度に比例して胃の中の頻度が変わる場合は餌選好が変わったとは言えない。
捕食者によっては、単一の餌だけを食べていてはすべての栄養素を摂取できないことがある。この場合、環境中の餌組成にかかわらず、胃の中の餌のメニューを豊かにしようとする傾向が生じる。結果的に少ない餌を一生懸命探す。このように少ない餌を好む場合を負の切り替え(negative switching)という。[]
逆に、多い方の餌が好まれることもある。2種類の被食者が別の場所に住んでいるときや、被食者によって姿や形が違う場合は、捕食者は同時に探すことができない。少なくとも、一方を探しているときは他方の種と遭遇しても発見しづらくなる。この場合、少ない餌を探すのは効率が悪い。そのときどきで数の多い餌を好むことを正の切り替え(positive switching)という。
一方の被食者の数が増えたとき、それを利用する天敵も増える。しかし必ずしも他方の被食者を狙う天敵が増えるとは限らない。天敵の大半は、今まで以上に増えた餌を狙い、他方の被食者を狙う天敵は減るかもしれない。逆に第2の被食者の数が増えたときには、そちらに天敵が集中し、第1の被食者の被食圧が下がる。したがって、捕食者が餌選好性を切り替える場合には、2種の被食者は互いに双利関係になり得る。我々はこれを巻き添え競争(apparent compeition)の対立概念という意味で、身代わり共生(apparent mutualism)と名付けた。[, ]英語の”apparent”は「見かけの」という意味である。つまり、一見競争がありそうなのだが実はないという印象を与える用語である。しかし実は逆である。一見関係のなさそうな種間の競争や共生関係のことである。
明らかな身代わり共生の実証例はまだ知られていない。しかし巻き添え競争の理論が示されてから実証例が報告されるまで約20年ほど経っているのだから、身代わり共生の実証にもあと20年近く待つ必要があるかもしれない。一般に、正の切り替えは被食者の共存を促し、被食者間の競争を緩めると考えられている。これは、捕食者の行動の変化を考えなければ説明できない、形質変化を通じた間接効果の典型である。
襲い分けによる捕食者の共存
行動を変えるのは捕食者だけではない。被食者もただ漫然と捕食者の餌食にされるのを待つのではなく、食われるのを避けようと努めている。草食動物だけでなく植物でも、被食回避形質の存在が知られている。発見されにくいような隠蔽色、まずい味にする化学防御、身を守るトゲや硬い皮膚、擬態、さらには群れて1個体当たりの被食率を下げたり、天敵の天敵を呼び寄せる外分泌物質(allelochemical)を出すものもいる。[,]
もっと素朴に、ときどき天敵がいないかあたりを見回したり、じっと隠れ家に身を潜めることもある。しかし草食動物も、四六時中隠れているわけにはいかない。自分自身も餌を探して食べたり、繁殖期には交尾相手を探したり、子育てをしなくてはいけない。ごく単純に採餌している間と隠れている間だけを考え、採餌中は天敵に対しても無防備で、隠れている間は安全と仮定する。
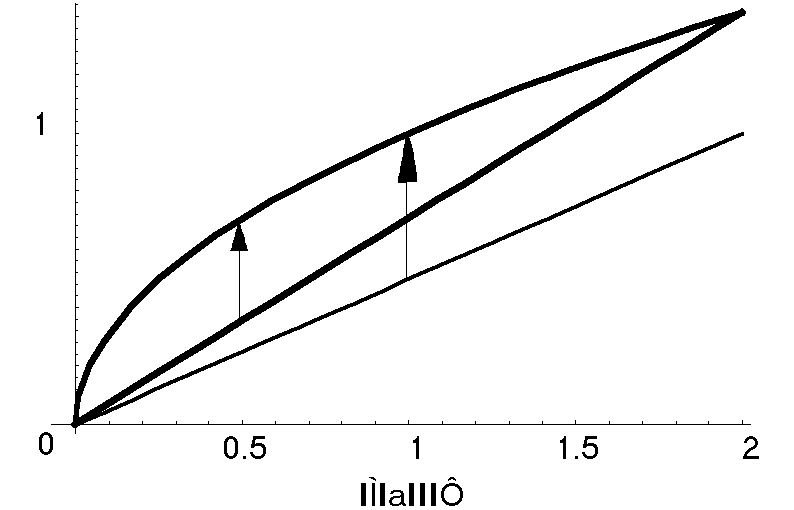
採餌時間(foraging time)をいくら延ばしても、2倍食べれば2倍子供を産むというものではない。おそらく、適応度(次世代に残す子供の期待数)は採餌時間に対して上に凸の増加関数になるはずである(図
1)。それに対して、草食動物が天敵に襲われる危険性は、草を食べる採餌時間が長いほどそれに比例して高くなるだろう。そうだとすれば、適応度Fは、たとえばF =
? cR - cPと表すことができる。ここでcは採餌時間、Rは被食者個体数(密度)、Pは天敵の個体数である。平方根は採餌による利益が摂食量cRの飽和関数であることを表している。
このとき、適応度Fを最大にする採餌時間cは図 1のように中間の採餌時間で適応度が最大になるはずである。上の式を微分すれば、Fが最大になる採餌時間はc=R/4P2と表すことができる。一般に採餌による利益が平方根でなく、f(cR)などと一般の関数であっても、最適採餌時間は天敵の数が増えるほど短くなることが理論的に証明できる。そして、実証的な報告例も知られている。[]
この草食動物の餌である草が多くなると採餌時間は長くなるだろうか、それとも短くなるだろうか。これは1990年代前半に論争の的となった。理論的には何とも言えないとする意見[]と、多くの場合、餌が多い方が早めに採餌を切り上げて身を隠すという意見が対立した。[]上記のようにf=
? cRと表せるときには、c=R/4P2は餌の量Rが多いほど長くなる。f=log(cR)の場合には餌の量Rによらず最適採餌時間は一定で、c=1/Pとなる。f=cR/(1+cR)の場合にはc=1/? PR - 1/Rであり、中間のRで採餌時間は最長になる。被食回避行動(形質)には、すべての天敵に有効な非特異的防御と、ある天敵にだけ有効な捕食者特異的防御(predator-specific defense)が考えられる。後者の好例がタンガニーカ湖の岩礁魚類群集に見られる。この湖にはカワスズメ科の魚が143種以上生息し、そのうち139種はこの湖の固有種であるという。カワスズメ科の魚の中には、他の魚の鱗を食べる鱗食魚が4種知られている。そのうち2種は同じ岩礁に共存し、両者がともに利用する「被食者」がたくさんいる。つまり、住み分けも食べ分けもしていない。両者は取り合い競争の関係にあるかというと、そうではないらしい。[]
2種は襲い方が異なる。一方(Perissodus microlepis)は底沿いに遠くから忍び寄って突進し、他方(P. straeleni)はひらひら舞いながら近寄って飛びつくという。[]襲い方が違うために被食者は同時に警戒できず、他方が近くにいるときの方が捕まえる確率が高くなると言う。さらに、それぞれの種に顎が右を向いた個体と左を向いた個体がいて、それぞれ被食者の左側面と右側面の鱗を狙い、右利きが多いときには被食者は左側面を警戒して右側面の防備を怠り、左利きの捕食者が有利になると言う。右利きと左利きは遺伝的に決まっている。その結果、右利きが多数派の世代と左利きが多数派の世代が4年周期で繰り返されていることが示唆された。[]4年という周期はこの魚の世代時間から理論的に説明できる。[]
環境が安定し、個体数変化もほとんどない熱帯の多様な魚類生態系の中にも、形質変化が引き起こす振動現象があることは、生態系が非定常であることの普遍性を示唆している。我々は、このように襲い方を変えて被食者の警戒を分散させて捕食者が共存する仕組みを「襲い分け」と名付けた。同じ餌を共有する2種の捕食者の関係も、必ずしも競争関係とは限らず、取り合い共生(exploitative mutualism)の関係になる場合がある。[、]
分別ある捕食者と乱獲問題
適応的な形質変化とは、必ずしも種の繁栄をもたらすとは限らない。これは利己的遺伝子(selfish gene)という用語とともに、生態学の常識になった。その好例は、先ほど説明した採餌時間である。草食者だけでなく、肉食者も四六時中餌を探しているわけではない。草食者が採餌時間を短くするのは天敵への備えだと説明したが、その説明は最高位の捕食者には当てはまらない。
以前は、捕食者の採餌時間が長すぎると被食者を乱獲し、かえって捕食者の個体数が減ってしまうからだと説明されていた。この説明は誤っているが、この問題を考えるために以下のような簡単な数理模型を考える。
dR/dt = (r - R/K) R - cNR
dN/dt = (- m +
? cR) Nただし、RとNはそれぞれ被食者と捕食者の個体数(時間tとともに変わる変数)、rとKは被食者の内的自然増加率と環境容量、cとmはそれぞれ捕食者の平均採餌時間と死亡率である。採餌量cRが多いほど利益が飽和することを、前節と同じく平方根ヨ cRで表した。このとき、定常個体数はdR/dt=0とdN/dt=0の連立方程式を解いてR=m2/c, N=(cK-m2)r/c2Kとなる。
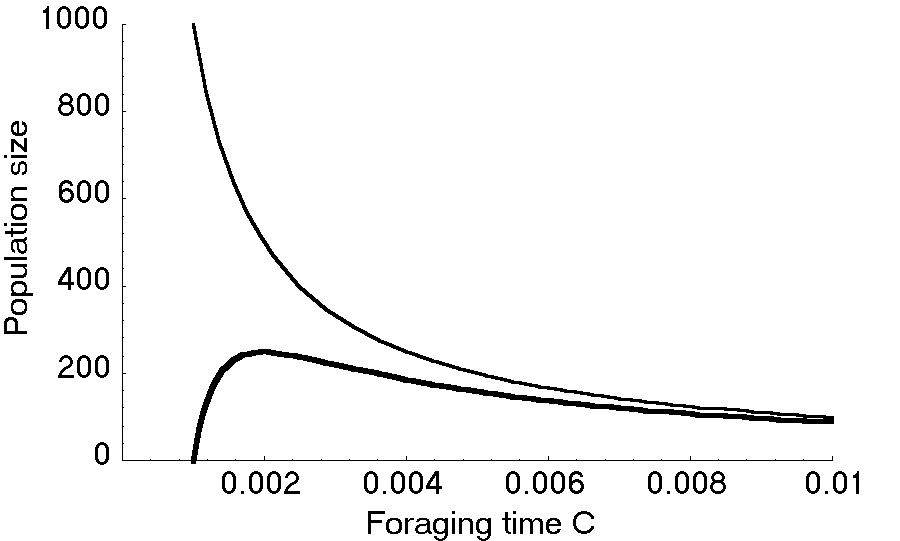
この数理模型では、第一に、被食者の定常個体数は捕食者の採餌時間が長いほど減ることがわかる。漁業でも、漁船の数や操業日数が増えたり、漁具効率が高くなると魚が減る。これを乱獲という。全く採餌しなければ(c<m2/K)捕食者は存続できないが、採餌時間が長すぎても捕食者の個体数は減り、c=2m2/Kのときが個体数最大でKr/4m2となる(図 2)。
これを「分別ある捕食者(prudent predator)」という。しかし生物は種全体が繁栄するように進化するのではない。自分の子孫が増えるような形質が進化すると考えられている。漁業でも、乱獲を防ぐのは自由参入の漁場では難しい。それと同じく、捕食者は他の個体より採餌時間を増やす方が他個体より適応度が高くなる。この理論では捕食者が採餌時間を縮めるのは説明できない。[]
採餌時間に限りがあるのは、採餌に労力がかかるからと考えられている。その労力は採餌時間に比例するか、満腹時になるほどさらに疲れるだろう。だから、天敵がいない最上位捕食者でも採餌時間は有限である。
外圧がもたらす繁栄
しかし、天敵や労力を考えても、採餌時間は維持できる個体数が最大になる状態ではなく、それより長い乱獲状態(過当競争)に落ち着く。そのとき、天敵の数が増えるとどうなるだろうか。
我々は、天敵が増えると被食者が減ると考えがちである。両者の形質がいつも変わらないなら、そしてこの2種以外の状況が変わらないなら、その直感は正しい。けれども、第3の種を通じた間接効果や、形質変化を通じた間接効果を考えると、この直感は正しいとは限らない。
植物と草食者、それを食べる肉食者を考える。それぞれの個体数をR, N, Pとする。ここでは簡単のために肉食者の数は一定とし、RとNの個体数変化が以下の数理模型で表せるとする。
dR/dt = (r - R/K) R - cNR
dN/dt = (- m +
? cR - cP) Nこれは前節の式に天敵に食べられる項cPをつけただけである。草食者が肉食者に食べられる頻度は、草食者自身の採餌時間に比例すると仮定した。草食者にとって最適な採餌時間は、
ある被食者密度Rに対して(- m + ? cR - cP)が最大になるc=R/4P2である。これをdN/dt=0の右辺に代入して解くと、植物の定常個体数R=4mPとなる。これをdR/dt=0の右辺に代入すると草食者の定常個体数N=rP/m-4P2r/Kとなる。驚くべきことに、肉食者Pがいないときよりも少しいたときの方が、草食者の個体数が増える(拙著[]113頁の図を参照)。あまりたくさん天敵がいるとさすがに減るが、天敵が多い方が草食者の数が減るとは限らないのである。これは直感的には、天敵の存在によって過当競争をやめ、結果的に分別ある捕食者に近づくためだと考えられる。
漁業では魚が魚自身の餌を枯渇させたり、漁船から逃げるという状況にはないかもしれないが、大発生したシカのような野生動物では、狩猟者やオオカミの導入がシカを減らすとは限らない。
このように、形質変化を通じた間接効果は、個体数変化にも思わぬ効果を及ぼす。私たちが生態系を理解し、人為的影響を予測する上で、しばしば今までの直感に反する影響が生じることを覚悟しなくてはならない。そのためには新たな直感を磨く必要がある。生態系を非定常系としてとらえることと、利己的遺伝子という用語に象徴される進化生態学の視点は、その重要な糸口になると思われる。
図の説明
図 1 採餌時間と採餌による利益、及び被食率の関係。採餌利益と被食率の差が最大になるとき(矢印)が最適な採餌時間であり、天敵の数が多いとき(太線)には少ないとき(細線)より短い。
参考文献
[] 伊藤嘉昭、山村則男、嶋田正和::動物生態学、蒼樹書房 (1992)
[] 松田裕之:「共生」とは何か-搾取と競争をこえた生物どうしの第三の関係, 現代書館: (1995)
[] M
üller C. B. and Godfray H. C. J.: Apparent competition between two aphid species. J. Anim. Ecol. 66, 57-64 (1997)[] 松田裕之: 種間相互作用と共進化, さまざまな共生(大串隆之編)平凡社、159-182 (1992)
[] Abrams, P. A., Menge B. A., Mittelbach, G. G., Spiller, D. A. and Yodzis P.: The role of indirect effects in food webs. In: Food webs: Interaction of patterns and dynamics (eds G. Polis and K. O. Winemiller), Chapman and Hall. pp.371-395 (1995)
[] 松田裕之: 野生生物の餌料選択と捕食者-被食者系の安定性, 中央水研研報, 第2号:53-64 (1991)
[] Abrams, P.A. and Matsuda, H.: Positive indirect effects between prey species that share predators. Ecology 77, 610-616 (1996)
[] 森主一: 動物の生態, 京都大学出版会 (1997)
[] 松田裕之: 覚りの数理生態学(3) 動物にはどれだけの採餌時間がいるか?, 遺伝, 48(3), 63-67 (1994)
[] Abrams, P. A.: Should prey overestimate the risk of predation? Am.Nat. 144, 317-328(1994)
[] Bouskila, A. and Blumstein, D. T.: Rules of thumb for predation hazard assessment: predictions from a dynamical model. Am. Nat. 139, 161-176 (1992)
[] 堀道雄: 多様な種間関係と多種共存、タンガニイカ湖の魚たち(堀道雄編)平凡社、pp.120-142 (1993)
[] Hori, M.: Frequency-dependent natural selection in the handedness of scale-eating cichlid fish. Science 260, 216-219 (1993)
[] Takahashi, S. and Hori, M.: Unstable evolutionarily stable strategy and oscillation -- a model on lateral asymmetry in scale-eating cichlids, Am. Nat. 144, 1001-1020 (1994)